Archives
- 2026-04
- 2026-03
- 2026-02
- 2026-01
- 2025-12
- 2025-11
- 2025-10
- 2025-09
- 2025-03
- 2025-02
- 2025-01
- 2024-12
- 2024-11
- 2024-10
- 2024-09
- 2024-08
- 2024-07
- 2024-06
- 2024-05
- 2024-04
- 2024-03
- 2024-02
- 2024-01
- 2023-12
- 2023-11
- 2023-10
- 2023-09
- 2023-08
- 2023-07
- 2023-06
- 2023-05
- 2023-04
- 2023-03
- 2023-02
- 2023-01
- 2022-12
- 2022-11
- 2022-10
- 2022-09
- 2022-08
- 2022-07
- 2022-06
- 2022-05
- 2022-04
- 2022-03
- 2022-02
- 2022-01
- 2021-12
- 2021-11
- 2021-10
- 2021-09
- 2021-08
- 2021-07
- 2021-06
- 2021-05
- 2021-04
- 2021-03
- 2021-02
- 2021-01
- 2020-12
- 2020-11
- 2020-10
- 2020-09
- 2020-08
- 2020-07
- 2020-06
- 2020-05
- 2020-04
- 2020-03
- 2020-02
- 2020-01
- 2019-12
- 2019-11
- 2019-10
- 2019-09
- 2019-08
- 2019-07
- 2019-06
- 2019-05
- 2019-04
- 2018-07
-
In terms of its protease activity MME has a
2020-08-19
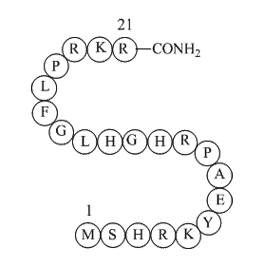
In terms of its protease activity, MME has a broad range of substrates being able to target glucagon, bradykinin, GLP1, and several other Aldicarb of circulating small molecules [25]. MME has been shown to target free insulin B-chain [32], although whether MME could target and degrade the insulin r
-
Fidaxomicin Dysfunction of the ubiquitin proteasome system c
2020-08-18
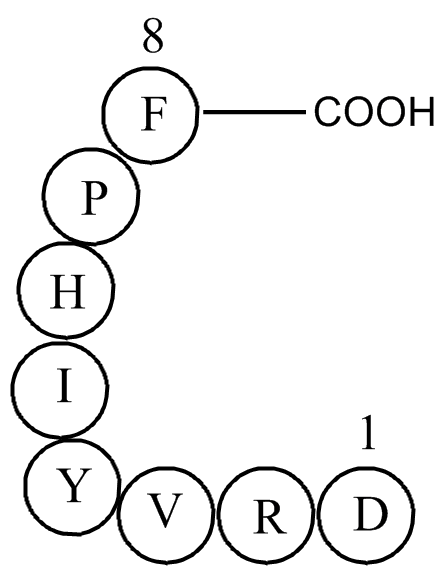
Dysfunction of the ubiquitin-proteasome system causes abnormal organelle morphology and impairs sperm formation and maturation steps resulting in male infertility [[7], [8], [9]]; this indicates that ubiquitination is essential for spermatogenesis and sperm function during fertilization. β-catenin i
-
Cobicistat receptor Eleutherococcus senticosus Rupt Maxim Ma
2020-08-18
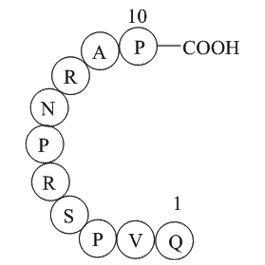
Eleutherococcus senticosus (Rupt. & Maxim.) Maxim is a shrub belonging to the Araliaceae, which is commonly distributed in China, Korea, Japan and Russia. It has been traditionally used as folk medicine for the treatment of rheumatism, diabetes, and hepatitis (Nan et al., 2004). Recent phytochemical
-
One possible explanation for differences
2020-08-18
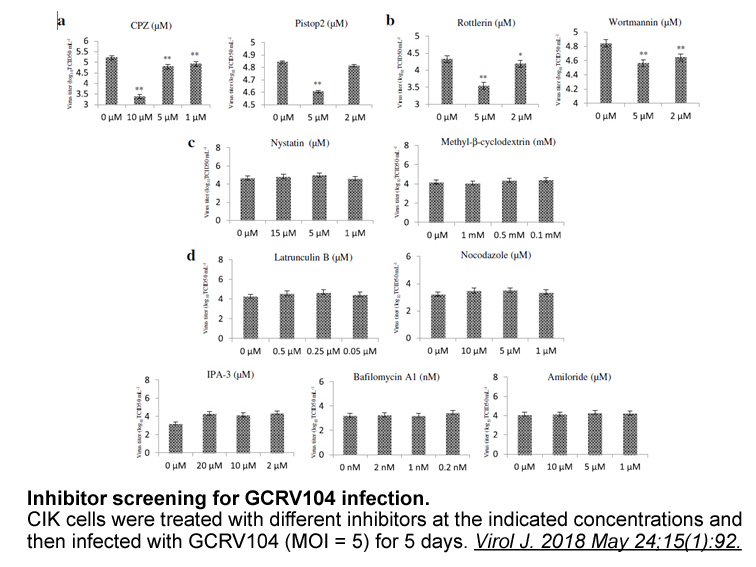
One possible explanation for differences in the binding ability of monomeric versus dimeric forms of DDR2 ECD to collagen could be that the monomeric form only binds to the primary GVMGFO site, whereas dimeric (and oligomeric) DDR2 ECD binds to additional sites on the collagen triple-helical molecul
-
In recent years the use of liquid chromatography
2020-08-18
In recent years, the use of liquid chromatography/tandem mass spectrometry (LC–MS/MS) has become a preferred approach to measure drugs and metabolites in biological fluids due to its very high selectivity and sensitivity [17], [18], [19], [20], [21], [22], [23], [25], [30]. The great sensitivity of
-
Perhexiline maleate CRF binds to CRF and CRF
2020-08-18
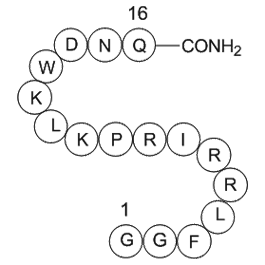
CRF binds to CRF1 and CRF2 receptor subtypes, triggering downstream cascades and leading to stimulatory G protein (GsP) activation. Once activated, GsP undergoes a structural change provoking the separation of the subunits Gα and Gβγ. The Gα subunit induces the production of the second messenger cAM
-
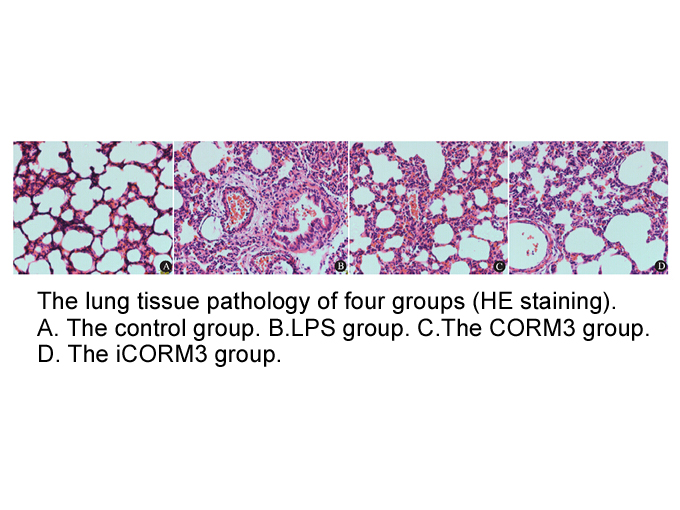
br Conclusion In this work experiments were
2020-08-18
Conclusion In this work, experiments were carried out to purify the protein with COX activity, using an anti-mouse COX-2 pAb and a COX activity assay as tools for monitoring the protein responsible for this activity in the parasite. Gp63 enrichment through the affinity column and identification b
-
MG 149 mg The recombinant Scl collagen system has shown
2020-08-18
The recombinant Scl2 collagen system has shown capability as a biomaterial as well because of its adaptability and scalability. Scl2 was functionalized to crosslink into a hydrogel without disrupting its triple helix [130]. The Scl2–hydrogel crosslinking also did not disrupt cell adhesion and integr
-
Impulsive burying and freezing behaviors in response
2020-08-18
Impulsive burying and freezing behaviors in response to aversive stimuli may represent attempts to avoid novel and/or aversive stimuli. These behaviors can be assessed utilizing the marble burying and conditioned fear tasks, respectively. For example, Wistar rats in proestrus and ovariectomized rats
-
In summary we have identified a series
2020-08-18
In summary, we have identified a series of amides as ERRγ agonists with suitable potency for further development. Our SAR studies of this chemotype identified SR19881 as the most potent full agonist of ERRγ with an EC=0.39 μM in a binding assay and an EC=4.7 μM in a cell-based assay. SR19881 was als
-
is one of the few parasites that can be
2020-08-18
is one of the few parasites that can be encysted and excysted and we tested the localization and role of HA-tagged gEno in both differentiations, using a recently developed integrated vector that introduces a C-terminal triple HA tag under the selection marker puromycin . The localization of HA-ta
-
In cancer interactions between the transformed cancer cells
2020-08-17

In cancer, interactions between the transformed cancer Atovaquone and other cell types recruited to the tumor are important. Tumor associated macrophages provide cancer cells with a suitable low-grade inflammatory milieu including growth promoting factors. Our results suggest a novel pro-tumorigenic
-
ARA metabolites which are produced through cytochrome P CYP
2020-08-17
ARA metabolites, which are produced through cytochrome P450 (CYP450) enzymes influence cardiovascular homeostasis. Human CYP4A11 and CYP1A2 metabolize ARA to 20-hydroxyecostearonic 98 receptor (20-HETE) [11], which is a vasoconstrictor [12], platelet activator [13] and naturetic in the kidney [14].
-
hUcn II influence on MAP is mediated by the
2020-08-17
hUcn II influence on MAP is mediated by the activation of CRF2 receptors. In vitro binding studies established that hUcn II is a selective ligand for CRF2 receptors with a slightly higher affinities for CRF2β compared with CRF2α and low or no affinity for CRF1 receptor [13], [19], [24]. Recent in vi
-
paroxetine hydrochloride In we studied normal rational varie
2020-08-17
In [1] we studied normal, rational -varieties X of complexity one, where the latter means that X comes with an effective torus action such that holds. We showed that for affine X with and at most log terminal singularities, the iteration of Cox rings is possible. In the present article, we charac
16175 records 828/1079 page Previous Next First page 上5页 826827828829830 下5页 Last page